The Velvet Touch of Slime
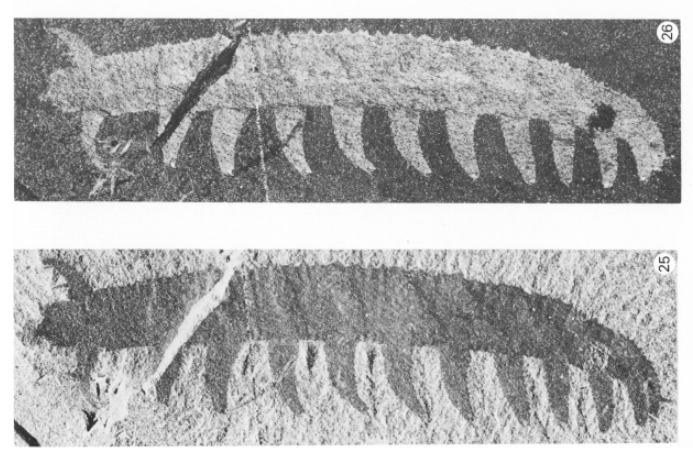
The first time I photographed this unusual slug/worm-like critter was during the construction phase of our Nectandra Cloud Forest Preserve in 2005. The 7.5 cm (3in) creature was slowly ambling around the wet algal carpet. It was brick red, with diamond shaped dorsal markings, purplish antennae, and a slightly flattened soft body, girdled with fine rows of “nubbins” on its skins (magnified view in lower photo B) and feet. It moved on two rows of 30 stubby, unarticulated, soft appendages. What was it? I was stymied. It had no exoskeleton, so couldn’t be a centipede or millipede. It couldn’t be a caterpillar because caterpillars have only 8 pairs of legs. It couldn’t be an annelid or slugs, because true worms or slugs have no legs.


I soon learned that it was commonly known as a velvet worm. It belonged in its own phylum Onychophora, named for their clawed feet. Each stubby, retractable foot is a hollow structure covering several sets of sharp claws that are stacked as in Russian dolls. The outer set is shed successively during each molting. The claws are for positioning and traction.
Onychophorans are a group of enigmatic organisms believed to be rare. True enough, I had to wait a full decade to see my second photo subject. In between those years, I only knew it by its common name. Few live specimens had been studied globally at the time and their family tree was being hotly debated among biologists.
Almost another decade went by. We still know little of the onychophorans because they are shy and infrequently seen. They are mostly nocturnal, although the two velvet worms I photographed appeared both during the day. What little we do know hints of the importance of the velvet worm’s placement in evolution. Interest revived recently among a handful of evolutionary biologists (mainly in Europe and Costa Rica) to learn about the velvet worms phylogenetic relationships to their nearest relatives — the arthropods (insects, arachnids, crustaceans), which constitute 80% of modern earth’s fauna.
What do we know about the ancestry of velvet worms?
They are from a very ancient lineage, and are often called living fossils having survived the multiple mass extinctions since the Cambrian. The earliest known fossils were no less than 500 millions years old. Tiny (≤ 3 cm) ancestors of the velvet worms, remarkably well preserved for soft-tissue organisms, were found in the Canadian Burgess Shale (Figs. C-E) These Cambrian ancestors, informally grouped as lobopods (Greek for blunt feet) shared stubby legs with two other ancient groups — the ancestral tardigrades (water bears) and arthropods (insects, spiders, centi/millipedes, crustaceans).

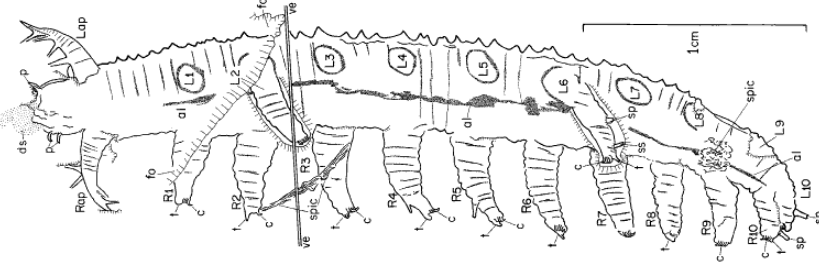
Anatomically, the onychophorans share many features with not just the tardigrades and arthropods, but also worms and nematodes. With no definitive identifying morphological traits, the phylogeny of the velvet worms languished for years, until the advent of DNA analysis and other non-destructive imaging technologies to study the 100 million yr old Burmese amber samples (Fig. G). We are finally beginning to glimpse into the lives of these curious organisms.
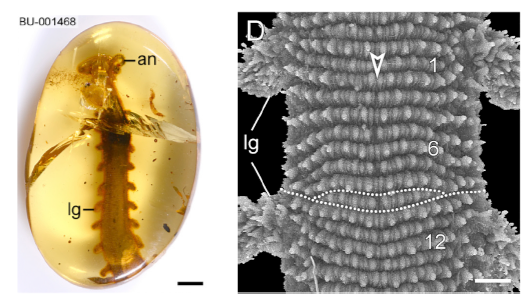
http://dx.doi.org/10.1016/j.cub.2016.07.023
Where do the onychophorans live
Without an exoskeleton, the terrestrial velvet worms are susceptible to desiccation. They are found in dark, damp, rotting plant litter or crevices in both the tropical and temperate wet forests. Their low density and infrequent sightings have hampered investigations into their biology, habitats, ecology and behavior. While we know about 200 species worldwide, most of the known natural history of the velvet worms comes from a period of interest between 1826 -1900. The more recent renewed attention and information indicate that there may be many more species yet to be discovered.
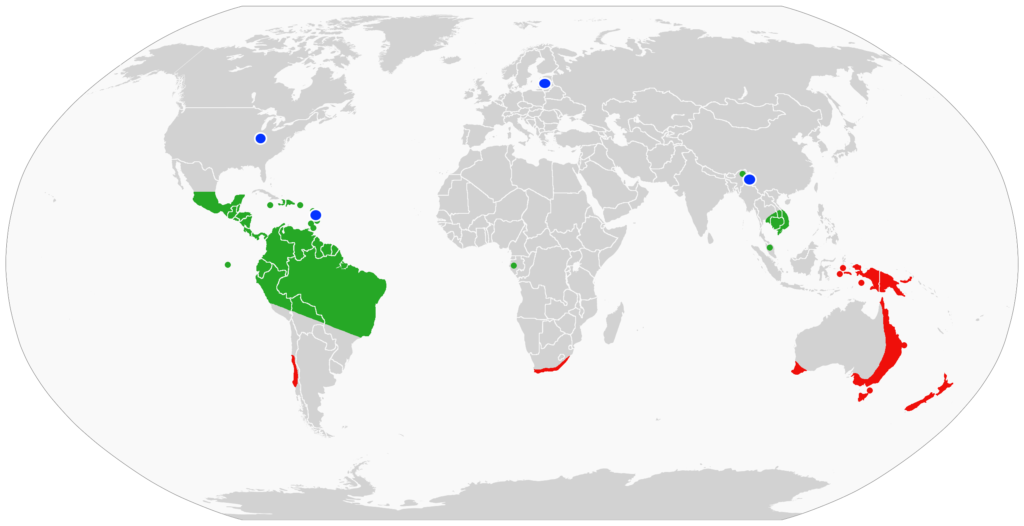
The largest number, 74 described species, of velvet worms is in Australia. A sampling of the Australian onychophorans show an array of resplendent velvety dorsal cuticles (Fig. I).

Unusual biology of the Onychophorans
The biology of velvet worms is nothing short of extraordinary.
Mating — Onychophorans reproduce sexually (except for one known hermaphroditic species). Actual mating has only been observed for 2 Australian and 1 Costa Rican species, with different variations on the mating positions.
In the Costa Rican species, the male and female join their abdominal ends for the insemination through the female vagina — the direct approach.
In one Australian species, the male deposits a packet of sperms (spermatophore) on the female skin on her back or sides. The spermatophore covering and the female skin are digested by the female’s enzymes to allow the sperms to migrate through the female body walls, and eventually through the ovary walls to fertilize the eggs — the indirect approach.
In the other variation, the male uses a special, ornate head structure to press the spermatophore directly into the female vagina (Fig. J). There is no information how the spermatophore is positioned on the male’s head in the first place. The female supports the male’s head with her hind claws while he positions the spermatophore. They remain joined for 15-30 min and could move as a joined pair, during which time the sperms are released into her vagina — the head-to-tail approach.

Thus far, there has been only one investigation into the fate of sperms after mating. Anatomically the female uterus is equipped with long-term sperm storage pouches containing sperms from multiple males, as well other accessory pouches containing decomposed sperms with mysterious ciliated funnels connecting the two.
Depending on the species, diverse reproduction of young is either by egg-laying or young-bearing. In the egg-laying species, females lay large yolk-filled eggs, while other species retain the eggs until they are ready to hatch. In the young bearing species, yolk-less eggs are nourished in the placenta. When the young are fully developed, the mother gives birth to her young similarly to placental mammals.
Feeding — They feed on small invertebrates and are voracious carnivores, hunt at dusk, subsisting on small invertebrates, worms and snails.
Hunting — The onychophorans most unique trait is their prey capturing technique. When feasible, they sneak up on the victim, poke it with its antennae to assess the prey’s size and risk. Alternatively, the velvet worm detects approaching prey through air movement. When ready to attack, it rears up its head, aims two large glands just under the antennae, then fires two gyrating jets of instant glue-like protein under high pressure. The fast acting, fast drying adhesive entangles the quarry within seconds. The velvet worm then spends many hours feasting afterward on the prey and consuming the nutritious slime. The latter is too nutritious to waste.
The following link is a short video showing how one physicist studies the fire-hosing technique in microscopic detail.